Следовые амины и рецептор следовых аминов 1: фармакология, нейрохимия и клинические последствия
Классические биогенные амины, в том числе дофамин (DA), норадреналин, адреналин, серотонин (5-HT) и гистамин, являются нейромедиаторами, которые играют важную роль в регулировании широкого спектра нейрофизиологических, поведенческих и когнитивных процессов, таких как двигательная активность, эмоции, мотивация, обучение и память. Изменения в аминоксидазной передаче широко распространены при различных неврологических расстройствах, в том числе при болезни Паркинсона (БП), шизофрении, синдроме дефицита внимания и гиперактивности (СДВГ), депрессии и наркотической зависимости. Соответственно, основной фармакологический подход к лечению таких состояний в основном основан на непосредственном изменении нейротрансмиссии этих классических моноаминов. Однако до настоящего времени был достигнут ограниченный успех в облегчении симптомов при этих расстройствах, при этом прямое манипулирование такими системами нейротрансмиттеров в некоторых случаях связано с нежелательными побочными эффектами. В этом отношении система DA является прототипом. Хроническое лечение леводопой по-прежнему превосходит другие методы лечения болезни Паркинсона, основанные на агонистах дофаминовых рецепторов, но приводит к долгосрочным, изнурительным двигательным осложнениям, таким как колебания активности и дискинезии. Аналогичным образом нейролептики, которые используются для лечения симптомов психозов и биполярного расстройства, могут вызывать экстрапирамидные побочные эффекты, характеризующиеся острой дистонией, акатизией, паркинсонизмом и поздней дискинезией. Кроме того, переносчик дофамина (DAT) уже давно используется для разработки заместительной терапии наркотической зависимости. Однако прогресс сдерживается потенциальной ответственностью за злоупотребление и неспецифическими эффектами, связанными с заменяющими лекарствами, основанными на фармакологии DAT, хотя сообщалось о многообещающих результатах с соединениями длительного действия. В ходе продолжающихся поисков более эффективных лекарств в нейропсихиатрии в последние годы всё больше внимания привлекает система передачи сигналов следовых аминов (TA) — вторичных аминов, тесно связанных с классическими биогенными аминами. ТА относятся к группе эндогенных аминов, нейробиологические функции которых оставались малоизученными до тех пор, пока в 2001 году две независимые исследовательские группы не открыли группу рецепторов, связанных с G-белком (GPCR), так называемые рецепторы, ассоциированные с таурином (TAAR). Исследования, проведённые за последние 15 лет, были сосредоточены в основном на TAAR1, поскольку это единственный представитель данного семейства рецепторов, который реагирует на ТА и филогенетически сохраняется в мозге млекопитающих. TAAR1 оказался важным модулятором основных биогенных аминов и начал набирать популярность как многообещающая мишень для разработки лекарств от различных нейропсихиатрических расстройств. В этом обзоре мы обобщаем последние достижения в понимании роли системы ТА в центральной нервной системе (ЦНС) млекопитающих, уделяя особое внимание её фармакологии и нейрохимии, а также важным функциям TAAR1 в патофизиологии и лечении нейропсихиатрических расстройств, связанных с нарушением регуляции аминоксилот.
Следовые амины
Тирамины, в том числе ρ-тирамин, м-тирамин, β-фенилэтиламин (β-PEA), м-октопамин, ρ-октопамин, триптамин и синефрин, представляют собой группу эндогенных аминов, которые содержатся как у беспозвоночных, так и у позвоночных. Хорошо известно, что у беспозвоночных следовые амины являются основными нейромедиаторами, а октапамин считается аналогом норадреналина в симпатической нервной системе позвоночных. Напротив, хотя существование следовых аминов (TA) в мозге и периферической нервной системе позвоночных было известно давно, их функции были в значительной степени неизвестны из-за очевидного отсутствия идентифицированных рецепторов, специфичных для ТА, что привело к их описанию как «ложных нейромедиаторов» . ТА структурно похожи на классические моноамины и имеют с ними общие пути биосинтеза и метаболизма. Первые исследования показали, что ТА оказывают симпатомиметическое действие на периферическую нервную систему позвоночных, связывая их с регуляцией кровяного давления и гомеостазом электролитов. Это понятие можно проследить до клинического наблюдения так называемой “сырной реакции”, гипертонического криза, испытываемого чувствительными пациентами, получавшими класс антидепрессантов - ингибиторов моноаминоксидазы (МАО) и подвергавшимися воздействию выдержанного сыра и других видов обработанных пищевых продуктов, обогащенных ρ-тирамином, образующимся в результате декарбоксилирования бактерий во время ферментации. Было показано, что периферический тирамин высвобождает эндогенный норадреналин из периферических запасов, что, в свою очередь, стимулирует адренергические нервы. Этот процесс отвечает за косвенное симпатическое действие.
В ЦНС ТА присутствуют в низких наномолярных концентрациях, которые в несколько сотен раз ниже, чем у классических нейромедиаторов, что может быть связано с их чрезвычайно высокой скоростью метаболизма и очень коротким периодом полураспада — около 30 секунд. Тираминсодержащие амины демонстрируют неоднородное распределение, которое в значительной степени соответствует классическим моноаминергическим проекционным путям, с повышенной экспрессией в нигростриарных и мезолимбических дофаминергических путях. Данные свидетельствуют о том, что некоторые тираминсодержащие амины, в том числе ρ-тирамин, β-PEA и триптамин, синтезируются в нигростриарных нейронах дофамина, а ρ-октопамин — в адренергических нейронах. Однако, как указывалось ранее, в течение длительного времени ТА считались просто метаболическими побочными продуктами других нейромедиаторов и не имели самостоятельного нейрофизиологического значения. В последующих исследованиях ТА были классифицированы как эндогенные нейромодуляторы, которые регулируют и сами регулируются совместно с другими нейромедиаторами. Действительно, многочисленные исследования продемонстрировали тесную функциональную взаимосвязь между ТА и классическими моноаминами, особенно дофамином. Во-первых, изменения в активности моноаминов способны влиять на уровень ТА. Например, было обнаружено, что снижение уровня β-PEA и ρ-тирамина в стриатуме увеличивает высвобождение дофамина, вызванное электрической стимуляцией чёрной субстанции, а ингибирование нейромедиатора дофамина приводит к повышенному накоплению β-PEA в стриатуме. В свою очередь, ТА, по-видимому, усиливают эффективность синаптической передачи, опосредованной этими моноаминами. Например, введение ингибиторов МАО-В, которые повышают уровень β-PEA выше физиологического диапазона, усиливает реакцию нейронов полосатого тела на дофамин и агонисты дофамина. Более того, ионофоретическое применение β-PEA вызывало усиление реакции нейронов коры головного мозга на норадреналин. Аналогичным образом, ионофоретическое введение ρ-тирамина, м-тирамина и β-PEA, применённые при слабых токах, усиливали реакцию нейронов коры головного мозга на дофамин. Аналогичным образом, применение октопамина при слабых ионофоретических токах усиливало как тормозящую, так и возбуждающую реакцию нейронов, опосредованную норадреналином. Влияние триптамина на нейротрансмиссию 5-НТ (серотонина) оказалось более сложным, поскольку наблюдалось как подавление, так и потенцирование 5-НТ-опосредованных нейрональных эффектов, что может быть объяснено двухфазными эффектами самого 5-НТ на активность нейронов коры. В совокупности эти результаты позволяют предположить, что ТА, вероятно, служат механизмом тонкой настройки, поддерживающим сбалансированный моноаминергический тонус за счёт реакции на колебания моноаминов, вызванные эндогенными или экзогенными факторами. Этот процесс может частично опосредоваться взаимодействием с определёнными рецепторами ТА.
Учитывая вышеупомянутую взаимную связь между ТА и моноаминами, неудивительно, что нарушение передачи сигналов ТА исторически ассоциировалось с широким спектром неврологических патологий, связанных с изменениями в работе моноаминов. Например, у детей с СДВГ был обнаружен пониженный уровень β-PEA в моче. Более того, в то время как у пациентов с депрессивными расстройствами был выявлен дефицит синтеза двух ТА, тирамина и октопамина, а также сниженная экскреция β-PEA с мочой, заместительная терапия PEA обеспечила долгосрочное облегчение депрессии у пациентов. Кроме того, у женщин с биполярным аффективным расстройством наблюдается очень высокая скорость выведения β-PEA с мочой, в то время как ингибиторы МАО, которые ещё больше увеличивают выведение β-PEA, усугубляют симптомы. Кроме того, хроническое лечение психогенными препаратами, диэтиламидом лизергиновой кислоты (ЛСД) и фенциклидином, приводило к повышению уровня мРНК декарбоксилазы ароматических L-аминокислот (AADC, фермента, катализирующего синтез β-PEA), что указывает на то, что избыточная выработка β-PEA может быть связана с патогенезом психотических синдромов. Действительно, было обнаружено, что уровень β-PEA в моче значительно повышен у параноидальных хронических шизофреников.
ТА также участвуют в действии психостимуляторов и, в более широком смысле, в формировании наркотической зависимости. Например, β-PEA имеет близкое структурное и фармакологическое сходство с центральным стимулятором амфетамином. В концентрациях, на несколько порядков превышающих его нормальный физиологический диапазон, β-PEA вызывал у грызунов и обезьян амфетаминоподобные эффекты, в том числе гиперактивность и стереотипное поведение. Эти данные позволили охарактеризовать β-PEA как «эндогенный амфетамин» мозга . Кроме того, активность β-PEA изменяется при экзогенном введении d-амфетамина. Острое введение d-амфетамина приводило к первоначальному снижению, а затем к повышению уровня β-PEA в мозге кроликов. Кроме того, хроническое введение амфетамина крысам снижало уровень мРНК AADC, что, в свою очередь, могло приводить к снижению активности β-PEA. С другой стороны, некоторые поведенческие эффекты амфетамина, по-видимому, зависят от уровня β-PEA, поскольку истощение запасов β-PEA в мозге блокирует стимулирующий моторику эффект d-амфетамина у мышей и кроликов.
Кроме того, появились доказательства того, что ТА могут влиять на вознаграждение (то есть субъективное ощущение удовольствия) и подкрепление (то есть усиление условной реакции на определённый стимул), что позволяет предположить участие ТА в неврологических адаптациях, лежащих в основе наркотической зависимости — хронического рецидивирующего синдрома, характеризующегося компульсивным употреблением наркотиков, неспособностью контролировать их приём и дисфорией при отсутствии доступа к наркотикам. В соответствии с предполагаемой ролью β-PEA как «эндогенного амфетамина», было показано, что он обладает свойствами, вызывающими привыкание, — определяющей характеристикой, лежащей в основе злоупотребления амфетамином и другими психомоторными стимуляторами. β-PEA также был так же эффективен, как и амфетамин, в формировании условного предпочтения места (т. е. процесса, при котором организм усваивает связь между действием наркотика и определённым местом или контекстом) у крыс и легко вводился собаками, у которых была устойчивая привычка (т. е. состоявшая из раннего приобретения и последующего сохранения) к самостоятельному введению амфетамина или кокаина. В другом исследовании высокие концентрации β-PEA в зависимости от дозы поддерживали реакцию у обезьян, которых ранее обучили самостоятельно принимать кокаин, а предварительное лечение ингибитором МАО-B, который задерживал деактивацию β-PEA, ещё больше повышало частоту реакций. Однако другие исследования выявили противоположное действие ТА на систему вознаграждения в мозге. У крыс, которые реагировали на пороговые уровни внутричерепной самостимуляции (ICSS) латерального гипоталамуса, антагонисты триптамина не только вызывали дозозависимое увеличение реакции при самостоятельном применении, но и усиливали стимулирующее действие амфетамина. И наоборот, систематическое применение триптамина снижало внутричерепную самостимуляцию (ICSS) как в медиальном ядре шва, так и в латеральном гипоталамусе. Эти результаты свидетельствуют о том, что обработка информации о вознаграждении в мозге подавляется посредством триптамин-опосредованного механизма. Кроме того, в отличие от условного предпочтения места, вызванного β-PEA, как отмечалось выше, триптамин вызывал условное отвращение к новому раствору сахарина у крыс, предполагающий негативное влияние на поведение, связанное с вознаграждением. Более того, хотя мало что известно о роли октопамина в вознаграждении мозга у позвоночных, функциональный вклад октопамина в обучение, связанное с вознаграждением, хорошо задокументирован у насекомых. Исследования, проведённые на дрозофилах, показывают, что тирамин необходим для развития чувствительности к кокаину. Считается, что это явление связано с теми же нейроадаптивными механизмами, которые лежат в основе тяги к наркотикам и рецидивов.
Несмотря на то, что эти результаты подчёркивают роль ТА в ключевых поведенческих и неврологических функциях, сигнальные механизмы и последующие молекулярные мишени, с которыми они связаны, оставались неясными до недавнего времени. В течение длительного времени преобладало мнение, что ТА действуют двумя способами. Во-первых, было высказано предположение, что ТА взаимодействуют с переносчиками в плазматической мембране, подавляя поглощение моноаминов и вызывая их отток посредством обратного транспорта, или препятствуют накоплению моноаминов в везикулах, вытесняя классические моноамины из их хранилища Во-вторых, ТА могут связываться с ещё не идентифицированными чувствительными к ТА сигнальными белками, расположенными на пре- или постсинаптических нейронах, содержащих рецепторы классических моноаминов, тем самым модулируя соответствующие внутриклеточные сигнальные пути. Более того, помимо изменения основных аминергических путей, было показано, что ТА модулируют нейронную передачу сигналов, опосредованную другими важными нейромедиаторами, такими как гамма-аминомасляная кислота и ацетилхолин, но функциональная значимость таких взаимодействий в настоящее время изучена недостаточно.
Идентификация семейства TAAR
Прогрессу в изучении нейробиологических функций ТА препятствовали трудности с выявлением их специфических рецепторов-мишеней и отсутствие селективных агонистов и антагонистов таких рецепторов. Хотя насыщаемые высокоаффинные сайты связывания, отличные от переносчиков и рецепторов аминов, были идентифицированы в головном мозге млекопитающих, именно в начале двадцать первого века две исследовательские группы независимо сообщили о клонировании и идентификации новое семейство GPCR млекопитающих. Такие рецепторы, в том числе несколько «сиротских» рецепторов, обладали необычайно высокой степенью гомологии последовательностей, и некоторые из них напрямую активировались ТА. Открытие рецепторов для TAAR подтвердило их роль в качестве настоящих нейромедиаторов, то есть молекул, способных напрямую запускать клеточные процессы, и привело к возобновлению интереса к ТА и их биологическим функциям. В последующих исследованиях Линдеманн и его коллеги предложили единую номенклатуру для этого недавно открытого семейства GPCR вместе с близкородственными рецепторами, назвав их рецепторами, ассоциированными с следами аминов (TAAR), признав тот факт, что некоторые из них не реагируют на следовые амины Дальнейшая работа той же группы исследователей позволила идентифицировать всех представителей этого семейства GPCR у крыс, мышей, шимпанзе и людей, продемонстрировав значительные различия в количестве генов рецепторов и доле псевдогенов у этих четырёх видов. У человека есть девять генов TAAR, включая три псевдогена; у шимпанзе — девять генов, включая шесть псевдогенов; у крыс и мышей — 19 и 16 генов, включая два и один псевдоген соответственно. Несмотря на эти значительные межвидовые различия, на основе филогенетических связей и сходства фармакофоров были выделены три подсемейства TAAR, которые оставались неизменными у всех четырёх видов. Три подсемейства включают TAAR 1–4, TAAR 5 и TAAR 6–9, при этом каждая подгруппа представлена как минимум одним функциональным геном TAAR. Неудивительно, что только два рецептора, активируемые TAs, а именно TAAR1 и TAAR4, относятся к первому подсемейству, что подтверждает функциональную основу классификации. В то время как TAAR1 чувствителен ко всем ТА, а ген TAAR1 филогенетически консервативен у всех изученных видов, включая человека, TAAR4 является псевдогеном в геноме человека, а TAAR4 у крыс реагирует только на β-PEA и тирамин, хотя и в гораздо меньшей степени, чем TAAR1. В результате TAAR1 получил наибольшее внимание за последнее десятилетие и является наиболее изученным рецептором этого класса.
Экспрессия, передача сигналов и фармакология TAAR1
TAAR1 связывается с Gαs G-белком и при стимуляции вызывает накопление внутриклеточного цАМФ посредством активации аденилатциклазы и стимулирует inwardly rectifying K+ каналы. Активация TAAR1 также может приводить к фосфорилированию PKA и PKC и усилению экспрессии факторов транскрипции CREB и NFAT. TAAR1 также передает сигнал через независимый от G-белка, β-аррестин2-зависимый путь, включающий сигнальный каскад протеинкиназы B (AKT)/гликогенсинтазы-3 (GSK-3) β, который играет важную роль во многих действиях, опосредованных дофамином. Среди ТА β-PEA и тирамин являются наиболее мощными активаторами TAAR1. β-PEA более эффективен, чем тирамин, в отношении TAAR1 человека и мыши, но не в отношении TAAR1 крысы. Поразительно, что в дополнение к ТА в качестве основных связывающих лигандов, TAAR1 также активируется огромным разнообразием эндогенных и экзогенных молекул, включая основные катехоламины, DA, норадреналин и серотонин и некоторые их метаболиты, амфетаминоподобные соединения, включая сам амфетамин, метамфетамин и 3,4-метилендиоксиметамфетамин (MDMA), производные спорыньи, включая ЛСД, и несколько адренергических лигандов, а также производные некоторых гормонов щитовидной железы. Исследования, направленные на определение распределения TAAR1 в организме млекопитающих, неизменно показывают, что мРНК или белок TAAR1 широко распространены и уникальны в центральной и периферической нервной системе человека, мыши, крысы и макаки-резуса. В головном мозге мРНК TAAR1 была обнаружена по всей лимбической системе и в областях, связанных с основными моноаминергическими путями, включая вентральную тегментальную область (VTA), черную субстанцию (SNr), голубое пятно, рафальное ядро, хвостатое ядро, путамен, прилежащее ядро (NAc), гиппокамп, гипоталамус и миндалину. Помимо подкорковых областей, TAAR1 также экспрессируется в корковых областях, особенно в пирамидных нейронах пятого слоя префронтальной коры (ПФК). TAAR1 преимущественно локализуется внутри клеток, диффузно экспрессируясь в периакрионе и вдоль аксональных отростков, а также в небольшом количестве в мембраносвязанных нейронах. Таким образом, было высказано предположение, что внутриклеточный TAAR1 может привлекать вспомогательный белок для транслокации к плазматической мембране или действительно может передавать сигнал внутри клетки, учитывая доступность нескольких эндогенных лигандов TAAR1 в цитоплазме и способность внутриклеточных рецепторов, сопряжённых с G-белком, оказывать влияние на нижестоящие процессы . Примечательно, что TAAR1 локализуется вместе с DAT в подмножестве нейронов дофамина и/или экспрессируется в нейронах, которые находятся в тесном контакте с нейронами, экспрессирующими DAT, в чёрной субстанции мышей и макак-резусов. Также есть данные, свидетельствующие о совместной экспрессии TAAR1 с переносчиком норадреналина в адренергических нейронах голубого пятна у макак-резусов, а также о совместной локализации с переносчиком 5-HT в серотонинергических нейронах дорсального ядра шва у мышей . Нейроанатомическое распределение TAAR1 по отношению к основным моноаминным системам позволяет предположить, что TAAR1 может регулировать моноаминергическую передачу посредством прямого взаимодействия с переносчиками моноаминов и пресинаптическими ауторецепторами, совместно экспрессируемыми с TAAR1 в отдельных нейронах, или посредством межклеточной коммуникации с близлежащими моноаминергическими нейронами. В следующем разделе мы рассмотрим доказательства, полученные в ходе различных исследований, в поддержку таких фундаментальных нейрохимических взаимодействий.
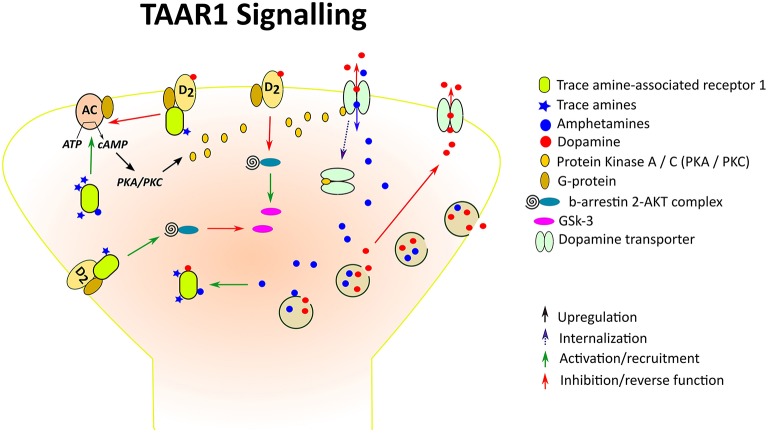
Функциональное взаимодействие TAAR1 с моноаминовыми системами мозга
Исследования с использованием гетерологичных систем экспрессии и синаптосом головного мозга выявили сложную трёхстороннюю взаимосвязь между TAAR1, переносчиками моноаминов и моноаминными авторецепторами in vitro. Во-первых, в таких препаратах in vitro активация TAAR1 агонистами, в том числе ТА, классическими биогенными аминами и препаратами амфетаминового ряда, была значительно усилена при совместной трансфекции TAAR1 с переносчиками моноаминов. Поскольку эти агонисты TAAR1 также являются субстратами для переносчиков моноаминов, была выдвинута гипотеза, что переносчики моноаминов могут служить проводниками для проникновения агонистов TAAR1 в синаптическое окончание, что позволяет активировать внутриклеточный TAAR1. В качестве альтернативы поглощение агониста, опосредованное переносчиком, может запускать перемещение TAAR1 в плазматическую мембрану. Кроме того, такие исследования in vitro показали, что активация TAAR1 функционально снижает активность переносчиков моноаминов. В клетках, одновременно трансфицированных TAAR1 и одним из основных аминных переносчиков, активация TAAR1 дофамином, норадреналином или 5-гидрокситриптамином приводила к функциональному ингибированию совместно экспрессируемого переносчика, снижая поглощение и увеличивая выведение связанного с ним нейромедиатора.
Более того, дополнительные доказательства свидетельствуют о том, что активация TAAR1 и моноаминовых ауторецепторов взаимно модулируют функцию переносчиков моноаминов посредством противоположных взаимодействий с аминотермическими передатчиками. В то время как активация TAAR1 способствует выведению моноаминов через их белки-переносчики, активация ауторецепторов приводит к увеличению поглощения классических биогенных аминов в стриатных и таламических синаптосомах обезьян и мышей дикого типа, при этом этот эффект отсутствует в синаптосомах мышей с нокаутом гена TAAR1 (KO) . Те же авторы показали, что при совместной трансфекции переносчиков дофамина, норадреналина и 5-HT с D2-рецепторами, α2A-рецепторами или 5-HT1B-рецепторами поглощение соответствующего аминного нейромедиатора значительно усиливалось. Аналогичным образом, норадреналин и серотонин также снижали удержание предварительно загруженного нейромедиатора в присутствии специфического антагониста ауторецепторов как у обезьян, так и у мышей дикого типа, но не у мышей с нокаутированным гена TAAR1, в синаптосомах. Зависимый от TAAR1 отток моноаминов объясняется обратным транспортом моноаминов через соответствующие переносчики, возникающим в результате опосредованного TAAR1 накопления цАМФ и фосфорилирования субстрата. В совокупности эти результаты указывают на то, что классические биогенные амины взаимодействуют как с TAAR1, так и с моноаминными ауторецепторами, регулируя функцию переносчиков. Таким образом, была предложена концепция пресинаптической рецепторной балансировки, согласно которой TAAR1 и моноаминные ауторецепторы уравновешивают активность моноаминов, при этом первые подавляют поглощение, а вторые способствуют ему.
Более поздние данные, полученные как в исследованиях in vitro, так и in vivo, указывают на прямое взаимодействие TAAR1 с моноаминными ауторецепторами, которое может лежать в основе ранее предложенной пресинаптической рецепторной балансировки. Во-первых, рецепторы TAAR1 и D2s при совместной экспрессии в клетках способны образовывать конститутивные гетеродимеры в плазматической мембране, что позволяет функционально регулировать эти два GPCR и/или другие клеточные субстраты. Действительно, уровень экспрессии TAAR1 как в общей массе, так и на мембране снижался при совместной экспрессии с D2s в тех же клетках. Такие взаимодействия между рецепторами TAAR1 и D2s частично опосредованы гетеродимеризацией рецепторов. Ранее было показано, что совместная трансфекция D2s, α2A, или α2B, или 5-HT1A, или 5-HT1B вместе с TAAR1 ослабляет вызванное активацией TAAR1 повышение внутриклеточного уровня цАМФ в ответ на дофамин, норадреналин или серотонин соответственно . Таким образом, благоприятное воздействие активации ауторецепторов на функцию переносчика моноаминов может быть результатом либо прямого усиления, либо подавления опосредованного TAAR1 блокирует функции переносчика. Важно отметить, что более поздние исследования показывают, что активация TAAR1 может также усиливать опосредованное D2s ингибирование передачи моноаминов — эффект, которого нет у мышей с нокаутированным геном TAAR1. Аналогичным образом, предыдущие исследования показали, что активация TAAR1 с помощью специфических агонистов повышает эффективность агонистов в отношении рецепторов 5-HT1A, в то время как избирательная блокада TAAR1 оказывает противоположный эффект. В совокупности эти результаты позволяют предположить, что стимуляция TAAR1 может оказывать двойное воздействие на моноаминергическую активность. В то время как прямое ингибирование TAAR1 переносчика моноаминов может привести к накоплению моноаминов во внеклеточной среде, опосредованное TAAR1 повышение активности моноаминовых ауторецепторов может привести к усилению функции переносчика и снижению передачи моноаминов. Однако важно помнить, что существуют разногласия по поводу наличия функциональных связей рецепторов in vivo, и следует соблюдать осторожность при оценке результатов на Гетеродимеризация in vitro и ее функциональные последствия. Например, в то время как гетероолигомеры рецепторов, связанных с G-белком, стали предметом многочисленных исследований из-за их потенциальной способности передавать сигналы иначе, чем рецепторы, из которых они состоят, что может иметь значение для разработки новых методов фармакотерапии, выявление таких комплексов in vivo было сложной задачей. Таким образом, необходимо тщательно изучить обоснованность этого подхода.
Таким образом, можно предположить, что относительная активация рецепторов TAAR1 и D2s эндогенными или экзогенными лигандами критически влияет на суммарную активность моноаминергических систем посредством ключевых воздействий на регуляцию переносчиков . В отличие от обычных биогенных аминов, которые активируют оба типа рецепторов, эндогенные ТА являются агонистами только для TAAR1. Следовательно, селективная стимуляция TAAR1 талидомидом или специфическими агонистами отличается от активации TAAR1 классическими моноаминами тем, что они вызывают ингибирующую модуляцию TAAR1 посредством совместной стимуляции с авторецепторами. Неудивительно, что in vitro совместная экспрессия моноаминовых ауторецепторов с TAAR1 ослабляла передачу сигналов TAAR1 в ответ на распространённые моноамины, но не на β-PEA. В соответствии с этими результатами, в то время как распространённые биогенные амины значительно усиливали поглощение в клетках, одновременно трансфицированных соответствующими моноаминными ауторецепторами и переносчиками, β-PEA этого не делал.
Кроме того, недавние исследования выявили регуляторное воздействие TAAR1 на постсинаптические рецепторы D2, что подчёркивает сложность молекулярных взаимодействий TAAR1 в моноаминовых синапсах. В то время как у мышей с нокаутированным геном TAAR1 нарушалось пресинаптическое аутоингибирование D2-рецепторов в полосатом теле, у них также наблюдалась повышенная экспрессия мРНК постсинаптических D2-рецепторов в полосатом теле и гиперактивность сигнального пути, опосредованного D2-рецепторами, G-белком/цАМФ-независимым, β-аррестином2-зависимым. Было показано, что активация рецептора D2 с помощью β-аррестина 2 приводит к дефосфорилированию AKT и его нижестоящей мишени GSK3β, что приводит к подавлению активности AKT и последующему усилению передачи сигналов GSK3β. Снижение фосфорилирования AKT с одновременным повышением уровня GSK3β связано с чрезмерной дофаминергической стимуляцией, вызванной либо косвенными агонистами дофамина, в том числе психостимуляторами, такими как амфетамин и кокаин, либо удалением переносчика дофамина (DAT). И наоборот, фармакологическое или генетическое ингибирование GSK3β отменяло стимулируемое дофамином поведение и снижало эффект психостимуляторов Таким образом, обнаружение того, что полосатые D2-рецепторы и связанный с ними сигнальный путь AKT / GSK3β повышены у мышей с нокаутом гена TAAR1 KO, предполагает, что активация TAAR1 может противодействовать передаче сигналов дофамина в постсинаптических участках, хотя этот эффект, вероятно, будет косвенным. Недавнее исследование показало, что образование гетеромерных комплексов между рецепторами TAAR1 и D2 не только снижает накопление цАМФ, стимулируемое TAAR1, но и смещает β-аррестин2 с активированных рецепторов D2 на активированный TAAR1. Таким образом, эти дополнительные данные свидетельствуют о том, что TAAR1 может снижать передачу дофамина не только за счёт усиления пресинаптического аутоингибирования, опосредованного D2-рецепторами, но и за счёт подавления постсинаптической передачи сигналов, опосредованной D2-рецепторами. В поддержку этой гипотезы можно привести тот факт, что мыши с дефицитом TAAR1 демонстрировали более высокую двигательную активность по сравнению с мышами дикого типа при воздействии квинпирола, агониста D2-подобных рецепторов, который, как известно, подавляет двигательную активность в низких дозах, стимулируя пресинаптические D2-подобные рецепторы, и усиливает двигательную активность в высоких дозах, активируя постсинаптические D2-подобные рецепторы. Это позволяет предположить, что дефицит TAAR1 может вызывать сверхчувствительность постсинаптических D2-подобных рецепторов.
В совокупности эти результаты свидетельствуют о том, что между TAAR1 и молекулярными мишенями моноаминов в пре- и постсинаптических участках происходят разнонаправленные взаимодействия. Однако знания, полученные на основе данных на клеточном и молекулярном уровнях, поднимают вопрос о конечном функциональном результате этих сложных взаимодействий и точной нейрофизиологической роли TAAR1. Изучение неврологических и поведенческих адаптаций, наблюдаемых у трансгенных мышей с модификациями TAAR1, даёт дополнительное представление об этом вопросе. По сравнению с мышами дикого типа, у мышей с нокаутированным геном TAAR1 не было выявлено различий в общих показателях здоровья, а также в общей двигательной активности, но наблюдались нарушения в подавлении акустического испуга перед импульсом, что является зависимой от дофамина реакцией, указывающей на нарушение сенсомоторной регуляции. Более того, у этих мышей с нокаутированным геном была повышена частота спонтанных разрядов и деполяризован мембранный потенциал покоя нейронов дофамина в вентральной области покрышки, увеличена частота спонтанных разрядов нейронов серотонина в дорсальном ядре шва и повышен уровень внеклеточного дофамина в прилежащем ядре. Кроме того, они продемонстрировали повышенную чувствительность к вызванной амфетамином двигательной активности и повышенный уровень внеклеточного дофамина, норадреналина и 5-гидрокситриптофана в полосатом теле, а также обусловленное метамфетамином предпочтение места. При воздействии МДМА («экстази») у мышей дикого типа наблюдалась дозозависимая двухфазная терморегуляционная реакция с ранней гипотермией, за которой следовала гипертермия, но у мышей с нокаутированным TAAR1 наблюдалась только длительная гипертермия, сопровождавшаяся повышенной чувствительностью к стимулируемой МДМА двигательной активности и высвобождению дофамина и серотонина в прилежащем ядре и дорсальном стриатуме, а также дофамина в лобной коре.
Напротив, у мышей, у которых была повышена экспрессия TAAR1, спонтанная двигательная активность не изменилась, но снизилась чувствительность к психомоторной активности, вызванной амфетамином, и высвобождению катехоламинов в прилежащем ядре. В совокупности эти результаты позволяют предположить, что TAAR1 может быть постоянно активным или тонически активироваться при повышении уровня эндогенных аминов, оказывая ингибирующее влияние на моноаминергическую нейротрансмиссию. Кроме того, эти данные свидетельствуют о том, что активация TAAR1 может снижать усиление моноаминергической передачи, вызванное стимуляторами двигательной активности. Активация TAAR1 амфетамином, метамфетамином и МДМА, которые сами являются агонистами TAAR1, может снижать нейрохимическое и поведенческое действие этих препаратов.
Исследования с использованием недавно разработанных высокоселективных агонистов и антагонистов TAAR1 предоставили более прямые доказательства того, что TAAR1 оказывает ингибирующее воздействие на передачу моноаминов. Селективный антагонист TAAR1, EPPTB, увеличил частоту разрядов дофаминовых нейронов в вентральной области покрышки у мышей, что согласуется с предположением о том, что TAAR1 постоянно активен или тонически активируется при повышении уровня аминов в окружающей среде, чтобы снизить активность дофамина. Более того, в то время как селективные полные агонисты TAAR1, RO5166017 и RO5256390, снижали частоту импульсов дофаминовых нейронов в вентральной области покрышки и серотониновых нейронов в дорсальном ядре шва, частичные агонисты, RO5203648 и RO5263397, повышали частоту импульсов этих же нейронов, что ещё раз указывает на высокую конститутивную активность или тоническую активацию TAAR1, которая служит для поддержания баланса передачи моноаминов. Эти результаты привели к предположению, что фармакологическая активация TAAR1 может служить средством модуляции изменённой передачи дофамина в патофизиологических состояниях. Последующие исследования подтвердили эту возможность. В то время как полный агонист RO5166017 снижал вызванное электричеством высвобождение дофамина как в дорсальном полосатом теле, так и в NAc (прилежащем ядре) на срезах мозга мышей, антагонист EPPTB повышал высвобождение дофамина в NAc . Кроме того, частичный агонист RO5203648 предотвращал вызванное кокаином высвобождение дофамина in vitro и временно ослаблял вызванное метамфетамином накопление дофамина в NAc in vivo, хотя и увеличивал частоту импульсов нейронов дофамина в вентральной области покрышки в обычных условиях. В поведенческих экспериментах как полные, так и частичные агонисты блокировали гиперподвижность, вызванную кокаином или амфетамином, у мышей дикого типа и мышей-мутантов с генетической делецией DAT. В совокупности эти результаты убедительно подтверждают роль TAAR1 как регулятора или стабилизатора системы дофамина.
Лиганды TAAR1 и их значение
Открытие TAAR1 в качестве распознаваемого сайта связывания для ТА не только подтвердило, что ТА являются настоящими нейромедиаторами, но и выявило потенциальный механизм, лежащий в основе различных нейропсихиатрических расстройств, связанных с дисфункцией системы ТА. Как обсуждалось выше, активация TAAR1 с помощью ТА может регулировать переносчики моноаминов и передачу моноаминов, что подтверждает, что TAAR1 является местом действия, через которое ТА контролируют нейрофизиологические процессы. Однако следует соблюдать осторожность при приписывании эффектов ТА рецептору TAAR1, поскольку у ТА могут быть и другие пути воздействия. Например, исследования показали, что β-PEA и тирамин обратимо снижают токи K+-каналов, опосредованные D2-ауторецепторами и управляемые G-белком, в нейронах дофамина среднего мозга мышей, воздействуя на другие участки, помимо TAAR1. Кроме того, некоторые побочные продукты метаболизма катехоламинов воздействуют на TAAR1. 3-метокситирамин, норметанефрин и метанефрин, мета-О-метильные метаболиты дофамина, норадрееналина и адреналина, соответственно, вызывают стимулированное TAAR1 повышение уровня ЦАМФ со значительно более высокой активностью, чем те, которые достигаются их исходным катехоламином (например, 3 МТ по сравнению с дофаминов в 2 раза; (±) NMN по сравнению с норадреналином (0,1)-4- раза; (±) MN по сравнению с адреналином — в 2,5 раза). Учитывая, что эти метаболиты обычно считаются биологически неактивными продуктами распада, обнаружение их высокой аффинности связывания с TAAR1 позволяет предположить, что их значение может быть выше, чем считалось ранее. Кроме того, было обнаружено, что прямые агонисты дофамина апоморфин и бромокриптин активируют TAAR1 in vitroж. Хотя степень, в которой их возможное взаимодействие с TAAR1 влияет на физиологические и поведенческие эффекты in vivo, остаётся спорным вопросом, недавние исследования показывают, что некоторые характерные стереотипные модели поведения, такие как лазание и облизывание, вызванные высокими дозами апоморфина, по крайней мере частично опосредованы TAAR1.
Учитывая структурное сходство между амфетаминоподобными соединениями и β-PEA, неудивительно, что амфетамин и некоторые производные амфетамина, в том числе метамфетамин, МДМА и 2-амино-1-[2,5-диметокси-4-йодфенил]-пропан (DOI), являются прямыми агонистами TAAR1. Постоянно сообщалось об их способности стимулировать выработку цАМФ у мышей, крыс, макак-резусов и человека TAAR1, экспрессирующийся в различных модельных клеточных системах, с активностью, сравнимой с β-PEA или тирамином, или немного более слабой, хотя также были отмечены видовые различия и изомерно-стереоселективность по агонистической активности и эффективности. Препараты из класса амфетаминов хорошо известны своей способностью повышать уровень моноаминов во внеклеточной жидкости головного мозга в основном за счёт того, что они действуют как субстраты переносчиков моноаминов, конкурируя с моноаминами за обратный захват, вызывая интернализацию переносчиков и способствуя опосредованному переносчиками оттоку (обратному транспорту). Они также препятствуют работе везикулярного переносчика моноаминов-2, истощая запасы моноаминов в везикулах и повышая доступность цитозольных моноаминов для обратного транспорта. Способность этих психостимуляторов высвобождать дофамин в больших количествах лежит в основе их усиливающих свойств и склонности к злоупотреблению. Величина высвобождения дофамина в вентральном стриатуме, вызванного амфетамином, положительно коррелирует с субъективным ощущением эйфории и желанием употребить наркотик, о котором сообщают люди. В связи с этим идентификация TAAR1 в качестве новой мишени для амфетамина и родственных ему препаратов раскрывает ранее неизвестный механизм действия стимулирующих препаратов и открывает новые возможности для разработки эффективных средств против зависимости. Исследования с использованием трансфицированных клеток, а также синаптосом стриатума мозга макак-резусов и мышей показали, что активация TAAR1 метамфетамином запускает ряд клеточных каскадов фосфорилирования, что приводит к снижению поглощения дофамина, усилению его оттока и интернализации DAT, что частично объясняет известные эффекты метамфетамина, связанные с высвобождением дофамина. Эти действия метамфетамина в отношении TAAR1 позволяют предположить, что фармакологическая модуляция TAAR1 может быть перспективным терапевтическим средством при зависимости от метамфетамина. Амфетамины сами по себе обладают терапевтической ценностью, хотя возможность злоупотребления ими сильно ограничивает их долгосрочную эффективность. Амфетамин используется для лечения СДВГ, который связан с нарушением моноаминергической модуляции функции префронтальной коры, а также нарколепсии и ожирения, которые также связаны с дефицитом функции дофамина. Улучшение когнитивных функций, противосонное и аноректическое действие амфетамина связывают с его способностью усиливать передачу моноаминов. Открытие того, что амфетаминоподобные соединения активируют TAAR1, повышает вероятность того, что TAAR1 может служить мишенью для терапевтического воздействия при этих неврологических и психопатологических состояниях, особенно с учётом того, что TAAR1 может напрямую контролировать функцию дофамина, как описано в предыдущих разделах.
Наконец, интересным открытием стало то, что несколько эффективных лигандов TAAR1 известны как антагонисты рецепторов биогенных аминов или ингибиторы переносчиков моноаминов. Например, адренергические или серотонинергические антагонисты фентоламин, толазолин и ципрогептадин, а также неконкурентные ингибиторы переносчика дофамина номифензин и 1-метил-4-фенил-1,2,3,6-тетрагидропиридин (MPTP) являются агонистами TAAR1. Следовательно, идентификация TAAR1 как места действия этих соединений может дополнить наше понимание их более известных антагонистических эффектов в отношении других рецепторов, связанных с G-белком, или белков-переносчиков. Напротив, кокаин, ингибитор DAT и мощный психомоторный стимулятор, проявлял низкое сродство к TAAR1.
TAAR1 и психические расстройства
Влияние TAAR1 на нейропсихиатрические расстройства широко распространено и описано в таблице 1. Болезнь Паркинсона характеризуется прогрессирующей дегенерацией дофаминергических клеток в нигростриарном пути и последующей потерей дофамина в полосатом теле. Лечение леводопой остается золотым стандартом фармакотерапии болезни Паркинсона благодаря ее способности частично восстанавливать уровень дофамина в полосатом теле. Однако длительное лечение леводопой вызывает инвалидизирующие побочные эффекты, такие как дискинезия и двигательные нарушения. Учитывая способность TAAR1 подавлять передачу дофамина, препараты, подавляющие активацию TAAR1, могут быть эффективны при лечении болезни Паркинсона. В поддержку этой гипотезы можно сказать, что эффективность леводопы в уменьшении симптомов паркинсонизма была выше у мышей с дефицитом TAAR1 по сравнению с мышами дикого типа . Более того, активация β-аррестина2 была связана со снижением уровня дискинезии, вызванной леводопой, без сопутствующего снижения терапевтической эффективности леводопы. Как обсуждалось выше, этот сигнальный путь регулируется TAAR1, что ещё больше укрепляет идею о том, что фармакотерапия на основе TAAR1 может быть эффективна при лечении болезни Паркинсона.
Несколько линий доказательств указывают на то, что TAAR1 играет роль в этиологии психоза. Во-первых, все гены TAAR человека плотно сгруппированы в узком участке хромосомы 6q23.1 размером 109 килобаз. , который близок или входит в число нескольких чувствительных локусов, которые были воспроизводимо связаны с шизофренией и биполярным аффективным расстройством в исследованиях связи или ассоциативных связей, предполагающих возможную причинную роль этого семейства рецепторов в этиологии психозов / полигенных нервно-психических расстройств. В соответствии с этим предположением, у мышей с нокаутированным геном TAAR1 наблюдалось повышение количества высокоаффинных рецепторов D2 в мозге и повышенная чувствительность к дофамину У пациентов с шизофренией во время психотического эпизода наблюдалось повышенное количество и чрезмерная стимуляция рецепторов D2. Кроме того, эффективность нейролептиков для захвата D2s была связана с клинической реакцией и экстрапирамидными побочными эффектам. Кроме того, у шизофреников наблюдается более сильная реакция на стимуляторы дофамина, такие как амфетамин и метилфенидат, по сравнению со здоровыми людьми. Эти результаты указывают на общие характеристики изменённой дофаминергической активности у мышей с дефицитом TAAR1 и у пациентов с шизофренией, что позволяет предположить, что аномальная функция TAAR1 может способствовать развитию психотических симптомов. Кроме того, в развитии шизофрении участвует нарушение регуляции сигнального пути β-аррестин2/AKT/GSK3β, и было показано, что широкий спектр клинически эффективных нейролептиков блокирует опосредованную рецептором D2 передачу сигналов β-аррестином2 и ингибирует активность GSK3β. Таким образом, обнаружение того, что стимуляция TAAR1 через β-аррестин2-зависимый путь приводит к подавлению GSK3β, является дополнительным подтверждением участия TAAR1 в развитии шизофрении и позволяет предположить, что препараты, усиливающие передачу сигналов β-аррестина2 через TAAR1, могут быть эффективным средством лечения этого заболевания. Кроме того, недавно появилась новая гипотеза D-нейронов, предполагающая участие системы TA в патогенезе шизофрении. D-нейроны — это немоноаминергические клетки полосатого тела, содержащие AADC, которые предположительно являются нейронами, вырабатывающими ТА. Согласно этой гипотезе, снижение пролиферации нервных стволовых клеток в субвентрикулярной зоне бокового желудочка приводит к уменьшению количества D-нейронов в полосатом теле и, следовательно, к недостаточному синтезу ТА. Последующее снижение стимуляции TAAR1 в конечном итоге приводит к увеличению высвобождения дофамина в NAc, а гиперстимуляция дофамина в стриатуме ещё больше подавляет пролиферацию нейронных стволовых клеток, усиливая гиперактивность дофамина. Однако это предположение в основном основано на наблюдении за уменьшением количества D-нейронов в стриатуме у пациентов с шизофренией и требует дальнейшей экспериментальной поддержки. Помимо дисфункциональной дофаминергической активности, сниженная глутаматергическая передача также была предложена в качестве причинного фактора при шизофрении, особенно в отношении негативных симптомов и когнитивных нарушений, связанных с заболеванием ). Антагонист NMDA-рецепторов, фенциклидин, широко используется в моделях шизофрении на грызунах для воспроизведения психотических симптомов, а некоторые атипичные нейролептики, как известно, модулируют NMDA-опосредованную нейротрансмиссию . Интересно, что избирательная активация TAAR1 как полными, так и частичными агонистами устраняет глутаматергическую гипофункцию, вызванную селективными антагонистами NMDA-рецепторов у животных дикого типа и у мышей-мутантов с дефицитом NMDA-рецепторов. Это позволяет предположить, что активация TAAR1 может усиливать глутаматергическую функцию. Более того, у мышей с нокаутированным TAAR1 белком наблюдался изменённый состав субъединиц корковых NMDA-рецепторов и нарушение регуляции NMDA-рецептор-зависимой синаптической функции, сопровождающееся повышенной персеверацией и импульсивностью. Более того, полная и частичная активация TAAR1 специфическими агонистами снижала импульсивное поведение у нормальных мышей, что указывает на благоприятную роль TAAR1 в передаче глутамата в коре головного мозга и связанных с этим поведенческих функциях. Кроме того, у мышей с нокаутированным TAAR1 наблюдались значительные нарушения сенсомоторной регуляции, измеряемые как ослабление подавления акустического испуга перед стимулом, что является известным поведенческим признаком шизофрении, регулируемым как дофаминергическими, так и глутаматергическими механизмами. Кроме того, селективные полные и частичные агонисты TAAR1, RO5256390 и RO5263397 соответственно, вызывали активацию мозга, схожую с той, что вызывается антипсихотическим препаратом оланзапином В совокупности эти данные убедительно указывают на то, что TAAR1 играет важную роль в этиологии и нейропатологии психоза, и подтверждают целесообразность изучения фармакологических препаратов на основе TAAR1 в качестве средств лечения этого заболевания.
Одним из наиболее хорошо задокументированных открытий в области фармакологии TAAR1 является способность этого рецептора регулировать нейрохимические и поведенческие реакции на психомоторные стимулирующие препараты, что привело к предположению, что селективные по TAAR1 соединения могут иметь терапевтическую ценность при лечении зависимости. Психостимулирующие препараты оказывают своё воздействие на ЦНС в основном за счёт сильных изменений в передаче дофамина и долгосрочных нейроадаптаций в дофаминергической системе. Действительно, тесное анатомическое и функциональное взаимодействие TAAR1 с системой дофамина и задокументированная роль TAAR1 в функциях вознаграждения и подкрепления в мозге согласуются с этой гипотезой. Более того, как обсуждалось в предыдущих разделах, исследования с участием трансгенных мышей и селективных агонистов TAAR1 убедительно продемонстрировали способность TAAR1 подавлять моноаминергическую реакцию на психомоторные стимуляторы. Несколько недавних исследований были направлены на изучение терапевтического потенциала агонистов TAAR1 на хорошо зарекомендовавших себя моделях наркотической зависимости на животных. Ревел и его коллеги предоставили первые доказательства того, что частичный агонист RO5203648 снижает вызванную кокаином двигательную активность и самовведение кокаина у крыс. Последующие исследования показали, что частичные и полные агонисты, RO5203648 и RO5256390 соответственно, снижали кривую зависимости от дозы при самовведении кокаина и предотвращали вызванное кокаином снижение порогов ICSS, что указывает на то, что частичная или полная активация TAAR1 блокирует усиливающие свойства кокаина. В соответствии с этими наблюдениями, другой частичный агонист TAAR1, RO5263397, снижал поведенческую сенсибилизацию, вызванную кокаином, и условное предпочтение кокаином места, где он был доступен. Важно отметить, что агонисты TAAR1 также предотвращали вызванный контекстом рецидив употребления кокаина и вызванное стимулами и кокаином возобновление поиска кокаина. В случае с метамфетамином частичные агонисты RO5203648 и RO5263397 снижали поведенческую сенсибилизацию, вызванную метамфетамином, и самовведение метамфетамина. RO5263397 также блокировал вызванное стимулами и наркотиками возобновление поиска метамфетамина. Примечательно, что в дозах, эффективных для ослабления поведения, связанного с употреблением наркотиков, такого как рецидив и возобновление употребления, эти агонисты не подавляли реакцию, вызванную естественным подкреплением, что исключает неспецифическое воздействие агонистов TAAR1 на общие двигательные и мотивационные функции. На самом деле, при использовании прогрессивного соотношения подкрепления частичный агонист RO5203648 снижал порог самоприменения кокаина, но усиливал реакцию на пищу (Pei et al., 2014), что указывает на явно различимые эффекты активации TAAR1 на реакцию на наркотик и пищу. Хотя механизмы, с помощью которых активация TAAR1 снижает стимулирующее воздействие на двигательную активность, до конца не изучены, ранее мы показали с помощью циклической вольтамперометрии с быстрым сканированием, что активация TAAR1 частичным агонистом RO5203648 предотвращает усиление передачи дофамина, вызванное кокаином, в прилежащем ядре без влияния на кинетику поглощения дофамина. Это позволяет предположить, что агонист TAAR1 ослабляет вызванное кокаином переполнение дофамином с помощью механизмов, отличных от прямого вмешательства в поглощение дофамина. В совокупности эти результаты показывают, что агонисты TAAR1 эффективно модулируют нейрохимические и поведенческие эффекты психомоторных стимуляторов и являются надёжной основой для разработки лекарств, нацеленных на TAAR1, для лечения зависимости от психостимуляторов. Кроме того, было доказано, что другие вещества, вызывающие зависимость, такие как опиаты, этанол, каннабиноиды и никотин, прямо или косвенно влияют на передачу дофамина. Хотя в психоактивном действии вызывающих привыкание веществ также участвуют процессы, не связанные с дофамином, способность TAAR1 модулировать передачу дофамина по-прежнему указывает на интригующую возможность того, что подход на основе TAAR сформирует новое поколение лекарств от зависимостей.
TAAR1 также может играть важную роль в эмоциональной регуляции, учитывая, что экспрессия TAAR1 повышена в миндалевидном теле, ключевой интегративной области, отвечающей за эмоции, эмоциональное поведение и мотивацию. Связь между дефицитом ТА и аффективными расстройствами, такими как депрессия, позволяет предположить, что активация TAAR1 может стабилизировать ненормальное настроение и эмоциональные колебания и способствовать действию антидепрессантов или анксиолитиков. В поддержку этой гипотезы было показано, что агонисты TAAR1 улучшают результаты в поведенческой парадигме с дифференциальным подкреплением низкой скорости у обезьян, сокращают время неподвижности в тесте на принудительное плавание у крыс и предотвращают вызванную стрессом гипертермию у мышей, демонстрируя таким образом антидепрессивные и анксиолитические свойства.
Последние данные свидетельствуют о том, что фармакологическое воздействие на TAAR1 может иметь более широкое применение и охватывать не только лечение заболеваний, но и укрепление психического здоровья. Было показано, что селективные лиганды TAAR1 улучшают когнитивные способности и повышают бодрствование. Несколько селективных полных и частичных агонистов TAAR1 повысили точность ответов в задаче на поиск предметов у обезьян и предотвратили вызванное фенциклидином снижение способности к переключению внимания у крыс, продемонстрировав прокогнитивные эффекты, в том числе повышение внимания, адаптивное торможение реакций и когнитивную гибкость. Кроме того, два селективных частичных агониста TAAR1 повышали бодрствование так же, как кофеин, но без побочных эффектов, связанных с кофеином, таких как повышенная двигательная активность и изменения температуры тела. В совокупности эти результаты позволяют предположить, что фармакология TAAR1 может иметь значение для улучшения когнитивных функций, контроля сна и психического здоровья в более широком контексте, связанном с производительностью труда и общим самочувствием.
Выводы
Новые открытия, о которых идёт речь в этом обзоре, раскрывают ключевую роль ТА и связанных с ними рецепторов в функциональной регуляции классических аминергических систем и подчёркивают важность TAAR1 как молекулярной мишени для разработки инновационных методов лечения неврологических заболеваний. Хотя физиологическое значение ТА не было полностью осознано до открытия семейства их рецепторов, ТА уже давно ассоциируются с классическими аминами. Нейрофармакологические исследования TAAR1, единственного представителя семейства TAAR, который филогенетически сохраняется у млекопитающих, значительно ускорились за последнее десятилетие, во многом благодаря разработке специальных инструментов для изучения его нейрофизиологических и поведенческих функций, в частности трансгенных моделей и прямых лигандов. TAAR1 взаимодействует с рецепторами DAT и DA D2, модулирует частоту импульсов нейронов DA и высвобождение DA, регулирует биохимические сигнальные каскады, запускаемые психостимуляторами, подобными амфетамину, и при избирательном удалении или активации обладает замечательной способностью влиять на ключевые модели поведения, связанные с неврологической дисфункцией. В результате растёт интерес к разработке методов лечения на основе TAAR1 для лечения ряда нервно-психических расстройств, включая болезнь Паркинсона, психоз, наркотическую зависимость и расстройства настроения. Кроме того, продемонстрированные агонистами TAAR1 свойства, способствующие пробуждению и улучшающие когнитивные способности, могут стать основой для разработки дополнительных методов лечения других форм дезадаптивного поведения. Однако наше понимание механизмов, лежащих в основе способности TAAR1 оказывать такое заметное регулирующее воздействие, остаётся недостаточным, и в будущем следует приложить усилия для полного изучения молекулярных путей и нижестоящих мишеней, которые задействуются TAAR1 в различных физиологических и нефизиологических условиях. Более того, разработка более совершенных исследовательских инструментов, таких как агонисты и антагонисты TAAR1, особенно последние, с улучшенным фармакокинетическим/динамическим профилем, позволит более эффективно оценивать TAAR1 как молекулярную мишень для разработки терапевтических средств в нейропсихиатрии. Несмотря на эти ограничения, убедительные доказательства теперь связывают TAs и TAAR1 с нейропсихиатрическими расстройствами, и по мере накопления новых знаний и появления более эффективных инструментов для изучения TAAR1 мы можем рассчитывать на новую эру фундаментальных и прикладных исследований в области нейробиологии.